 咨询热线: 176-001-76302
咨询热线: 176-001-76302 
高级灵长类胎盘雌激素合成的非经典途径
2023-05-08 10:27:19 点击数:通过卵巢雌激素合成经典途径可以看出,C19类固醇为雌激素合成的前体,17α-羟化酶(P450 c17 )则是转化C21类固醇激素(孕烯醇酮和孕酮)成为C19类固醇(脱氢表雄酮和雄烯二酮)的关键酶。如果组织中存在P450 c17 就可以利用胆固醇或孕激素为雌激素合成提供雄激素前体;如果组织中缺乏P450 c17 则不能利用胆固醇或孕激素为合成雌激素提供雄激素前体。高级灵长类胎盘缺乏P450 c17 ,因此不能利用胆固醇、孕烯醇酮和孕酮为雌激素合成提供雄激素前体。虽然高级灵长类胎盘不能利用孕激素合成雌激素,但高级灵长类胎盘含有高活性的芳香化酶,因此转化C19类固醇(脱氢表雄酮、雄烯二酮、睾丸酮)为雌激素的能力很强。胎盘C19雄激素的来源便成为灵长类胎盘雌激素合成的限速步骤。非灵长类动物胎盘存在17α-羟化酶,因此可利用胆固醇、孕烯醇酮和孕酮合成雌激素。所以,非灵长类动物的妊娠晚期,胎盘P450 c17 在胎儿来源的糖皮质激素的诱导下表达增加,导致孕激素向雌激素的转化增加,因此母体血液孕激素水平下降的同时雌激素水平升高。由于高级灵长类胎盘缺乏P450 c17 ,因此不存在孕激素向雌激素的转化,所以母体血液的孕激素和雌激素水平在妊娠晚期同时升高。
1)胎盘C19类固醇的来源:
1961年,Frandsen和Stakemann发现怀有无脑畸形儿的孕妇尿液的雌激素浓度只有正常孕妇的1/10。根据胎儿肾上腺皮质萎缩是无脑畸形儿的典型特征,他们认为胎儿肾上腺皮质可能是胎盘合成雌激素前体的主要来源。进一步的实验发现,胎盘可将注入母体血液循环的脱氢表雄酮硫酸酯(dehyroepiandrosterone sulfate,DHEAS)、脱氢表雄酮、雄烯二酮和睾丸酮迅速转化为雌激素,说明含有19个碳原子的雄性激素为胎盘雌激素合成的前体。实验还发现,孕妇血液的脱氢表雄酮硫酸酯含量很高,而且半衰期较长。妊娠第7周时,胎盘既具有转化脱氢表雄酮硫酸酯为雌二醇的能力;妊娠第30周时,至少25%的母体血液脱氢表雄酮硫酸酯被胎盘转化为雌激素,还有30%的脱氢表雄酮硫酸酯被母体肝脏转化为16α-羟基脱氢表雄酮硫酸酯。
妊娠期母体血液的DHEAS来源于何处呢?一般认为肾上腺为DHEAS的主要来源,但妊娠期母体肾上腺DHEAS的合成并不像胎盘雌激素的合成随孕期逐渐增加,说明母体肾上腺可能不是DHEAS的主要来源。由于怀有无脑畸形胎儿的孕妇胎盘雌激素的合成大大减少,脐带血DHEAS的浓度又很高,而且灌流入脐带动脉的DHEAS可被胎盘迅速转化为17β-雌二醇和雌酮,根据以上证据人们提出,胎儿肾上腺皮质可能是胎盘雌激素前体DHEAS的主要来源。事实上,母体和胎儿来源的DHEAS对胎盘雌二醇的合成同等重要。现在已经明确,妊娠晚期胎盘合成的17β-雌二醇中,50%来源于母体肾上腺提供的DHEAS,50%来源于胎儿肾上腺提供的DHEAS。值得指出的是,比较低级的灵长类如猴和狒狒妊娠期血液循环中的雌激素是以雌二醇为主,而高级灵长类如大猩猩、黑猩猩及人类妊娠期血液循环的雌激素主要是雌三醇。
一般认为雌三醇是由雌酮和雌二醇转化而来的,雌二醇C16位的羟基化是转化为雌三醇的关键一步。非孕期正常妇女尿液的雌三醇与雌酮和雌二醇之间的比例为1∶1。孕妇尿液中雌三醇与雌酮和雌二醇之间的比例为10∶1,接近妊娠足月时这一比例进一步升高,提示妊娠时雌二醇C16位的羟基化增多。但胎盘并无羟化雌二醇的功能,妊娠期雌三醇可能有以下三个来源:①由胎盘合成的雌酮和雌二醇先进入胎儿体内,由胎儿转化为雌三醇,然后再进入母体循环;②胎盘合成的雌酮进入胎儿体内,由胎儿转化为16α-雌酮后,再由胎盘还原为雌三醇;③胎儿和母体血液中存在一种可以作为胎盘合成雌三醇的前体的化合物即16α-羟基-C19-类固醇。虽然妊娠期这三条途径都存在,但以第三条合成途径为主。注射入母体血液的放射性核素标记物16α-羟基脱氢表雄酮及其硫酸酯可以被胎盘迅速转化为雌三醇。母体和胎儿血液中存在大量的16α-羟基脱氢表雄酮硫酸酯,可以为胎盘合成雌三醇提供足够的前体。虽然胎儿和母体都可以合成16α-羟基脱氢表雄酮硫酸酯,但由胎儿来源的16α-羟基脱氢表雄酮硫酸酯合成的雌三醇占胎盘雌三醇合成总量的90%,即胎盘雌三醇的合成前体主要来源于胎儿。现在已经明确,妊娠期胎盘雌三醇的合成主要途径如下:胎儿肾上腺首先合成DHEAS,然后DHEAS再在肝脏中羟化为16α-羟基脱氢表雄酮硫酸酯,后者随脐带血到达胎盘,在胎盘硫酸酯酶、芳香化酶的催化下转化为雌三醇(图5-3)。由此可见,妊娠期雌三醇是由胎儿、胎盘共同参与制造的,故把妊娠期雌三醇合成称为胎儿-胎盘单位。由于16α-羟基脱氢表雄酮硫酸酯主要来源于胎儿,因此胎盘雌三醇的合成量可以反映胎儿的健康状况,临床上检测母体血液雌三醇的含量可用来判断胎儿是否存活。
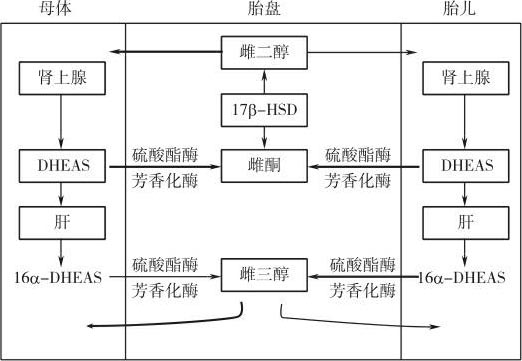
图5-3 妊娠期雌激素合成的胎儿-胎盘单位
由以上可以看出,DHEAS和16α-羟基脱氢表雄酮硫酸酯分别是胎盘合成雌二醇和雌三醇的前体,但无论DHEAS还是16α-羟基脱氢表雄酮硫酸酯都必须首先在胎盘脱去硫酸酯才能继续被芳香化酶转化为雌二醇或雌三醇。
2)硫酸酯酶(sulfatase):
胎盘合成雌激素的前体都是以硫酸酯的形式存在于血液的,由胎盘摄取后首先需要在硫酸酯酶的催化下脱去硫酸基。胎盘含有丰富的硫酸酯酶,硫酸酯酶主要存在于滋养层细胞的内质网内。当细胞滋养层细胞向合体滋养层细胞转化时,硫酸酯酶的表达增加。胎盘存在多种硫酸酯酶亚型,如芳基硫酸酯酶A、B、C,不同亚型硫酸酯酶的结构类似,他们都是由8个亚基组成的多聚体。胎盘合成的类固醇对硫酸酯酶具有负反馈性调节作用。先天性缺乏硫酸酯酶的孕妇,组织中硫酸酯化的胆固醇水平增高,导致男性胎儿出现干皮病(ichthyosis)。
3)芳香化酶(aromatase):
当DHEAS和16α-羟基脱氢表雄酮硫酸酯被脱去硫酸基团后,首先转化为雄烯二酮和雄烯三醇(5-androstene-3β,16α,17α-triol),然后再被芳香化酶转化为雌酮或雌三醇。芳香化酶属于细胞色素P-450酶类,它催化类固醇结构中C10位上甲基的裂解和A环的芳香化,从而将雄烯二酮、雄烯三酮分别转化为雌酮、雌三醇。芳香化酶主要存在于合体滋养层细胞的线粒体和微粒体中,芳香化酶基因已被克隆,长约有75kb,含有11个外显子,其中两个外显子不起转录作用。人类芳香化酶基因具有两个鲜明的特点:①第一个内含子很大,约有35kb,几乎占整个基因的1/2;②芳香化酶的基因存在多个转录启动位点,因此可表达出不同的mRNA及芳香化酶。芳香化酶基因与P450scc基因共同位于第15号染色体上,两种酶的基因共存于同一染色体的意义目前还不清楚。
4)17β-羟基类固醇脱氢酶:
雄烯二酮转化为雌酮后还需在17β-HSD的催化下,进一步转化为作用更强的17β-雌二醇。另外,脱氢表雄酮向雄烯二醇、雄烯二酮向睾丸酮的转化也需要17β-HSD的催化。17β-HSD存在于胎盘的微粒体内,它催化雌酮、脱氢表雄酮和雄烯二酮C17位的酮基还原为羟基成为雌二醇、雄烯二醇和睾丸酮。现在已有五型17β-HSD cDNA被克隆。根据克隆的先后,分别命名为17β-HSDⅠ、Ⅱ、Ⅲ、Ⅳ、Ⅴ型。Ⅰ型和Ⅱ型17β-HSD存在于胎盘组织,Ⅰ型酶催化雌酮向雌二醇的转化,Ⅱ型酶催化雌二醇向雌酮的转化。
(2)雌激素的作用:
妊娠期母体血液的雌激素中,以雌三醇的浓度为最高,但血液中的雌三醇95%是以结合状态存在的,雌酮和雌二醇与结合蛋白结合比例分别为75%和5%。因此,妊娠期的雌激素作用主要是由雌二醇产生的,雌酮的作用次之,雌三醇的作用最弱。雌酮可以在17β-HSD的作用下转化为作用更强的雌二醇发挥作用。雌激素为调节女性机体生理功能的重要激素,尤其对生殖系统和乳腺的调节更为重要。
- 上一篇:胎盘激素的作用方式和机制
- 下一篇:详解雌激素受体
